Стратегия генетического картирования и его роль в идентификации новых генов наследственных заболеваний. Принципы генетического картирования Генетические карты сцепления
Slide 1
Выполнила: Голубева Ю.В. 410гр
Slide 2
Одна из основных задач современной генетики
заключается в выяснении природы комплексных
признаков, к которым в частности относятся
многие распространенные болезни человека и
характеристики продуктивности
сельскохозяйственных животных. Стартовым
этапом на пути решения этого вопроса
является
Slide 3
Картирование генов -
Slide 4
Стратегические подходы
к картированию геномов
Slide 5
Стратегия прямой
генетики
Различия во времени появления,
необходимой методической базой и
спектре возможностей. Функция гена
известна хотя бы частично.
Slide 6
Функциональное
картирование
Основа - наличие некоторой информации о
биохимическом полиморфизме, лежащем в
основе того или иного наследственного
признака.
начинается с выделения в чистом виде
белкового продукта гена.
к нему по аминокислотной последовательности
подбирают вырожденные праймеры
проводят ПЦР-скрининг
Slide 7
Большинство генов, функция которых
была известна, уже клонированы и
локализованы.
Slide 8
Для большинства генов, которые
были локализованы, характерны
структурные аномалии (как
правило, это гены, ответственные за
наследственные заболевания
человека), что существенно
облегчает заключительную стадию
поиска гена - выделение и
локализацию гена.
Slide 9
Кандидатное
картирование
информация о функциональном
изменении недостаточно полна, чтобы
точно указать ген
Информации достаточна для того,
чтобы выдвинуть предположения о
возможных кандидатах либо по их
функции, либо по положению на
хромосоме
Slide 10
Общее:
при функциональном, и при
кандидатном подходе клонирование
гена, как правило, предшествует его
точной локализации в геноме
локализовать ген означает пройти путь
от его функции к локализации на
хромосоме (позиции)
Slide 11
Стратегия обратной
генетики
От хромосомной карты к функции
гена. Возникло благодаря появление в
конце 80-х годов множества
высокополиморфных ДНК-маркеров
Slide 12
Позиционное
картирование
локализация гена при отсутствии всякой
функциональной информации о нем
место гена на карте устанавливают по
результатам анализа его сцепления с
ранее локализованными генетическими
маркерами, далее исследуется уже
область генома рядом с маркером
Slide 13
Генетический маркёр
(genetic marker)
Ген, детерминирующий
отчетливо выраженный
фенотипический признак,
используемый для
генетического картирования
и индивидуальной
идентификации организмов
или клеток. Также в качестве
генетических маркеров
могут служить целые
(маркерные) хромосомы.
Slide 14
Минусы
ограничением позиционного
подхода является низкая
разрешающая способность
генетических карт - интервал между
двумя соседними маркерами, в
котором локализован ген, может
оказаться слишком велик и
недоступен физическому
картированию.
Slide 15
Картирование генов –
виды
Физическое картирование
Генетическое картирование
Цитогенетическое(цитологическое)
картирование
Slide 16
Физическое
картирование
обширная группа методов, позволяющая строить
карты генома (обычно их называют физическими)
высокого уровня разрешения и определять
расстояния между локализуемыми нуклеотидными
последовательностями с точностью от нескольких
десятков тысяч п.н. до одной нуклеотидной пары.
Пример: картирование
генов с помощью
хромосомных мутаций
Slide 17
Типы физического
картирования
рестрикционное картирование
RH-картирование
клонирование в YAC (от англ. yeast artificial
chromosome)
BAC (от англ. bacterial artificial
chromosome) в космидах, плазмидах и
других векторах и контиг-картирование на
их основе
секвенирование ДНК
Slide 18
В том случае, когда известна
последовательность ДНК интересующего
локуса, эту последовательность можно
использовать для гибридизации с
хромосомами in situ, и место гибридизации
будет однозначно указывать на локализацию
локуса в определенном районе определенной
же хромосомы
Slide 19
Генетическое
картирование
картирование, основанное
на методах классической
генетики - определении
групп сцепления, частоты
рекомбинации и
построении генетических
карт, где единицей
измерения служат
проценты рекомбинации
Slide 20
Первый ген человека
был локализован на
Х-хромосоме в 1911
г.
Первый аутосомный
ген - только в 1968 г
Slide 21
Генетическая карта
(genetic map
Схема взаимного
расположения генов на
хромосоме (в группе
сцепления) и их
распределения по
разным хромосомам,
как правило,
включающая данные об
относительном
удалении генов друг от
друга (генетические
расстояния).
Slide 22
Генетическая карта
американской норки
включает 127 генов
(черный текст) и 39
микросателлитных
последовательностей
(красным текст).
Разным цветом
выделены районы
хромосом норки
гомологичные
хромосомным.
Slide 23
Преимущества
большое число консервативных групп
сцепления
создание банков клеточных культур
для локализации вновь возникшей
мутации к настоящему моменту есть
набор маркерных генов для каждой
хромосомы.
Slide 24
Построение
генетической карты
Шаг 1: формирование групп
сцепления генов и исследование их
взаимного расположения(Скрещивания
проводятся до тех пор, пока не удастся выявить
сцепленное наследование анализируемой
мутации с маркерными мутациями какой-либо
хромосомы)
Шаг 2: подсчитывание расстояния
между исследуемым геном и уже
известными маркерными генами
Slide 25
Единицы измерения
Генетическое расстояние между линейно
расположенными генами, выраженно в процентах
рекомбинации -
Два гена на хромосоме
находятся на расстоянии 1
сМ, если вероятность
рекомбинации между ними
в процессе мейоза
составляет 1%.
Классический пример Моргана –
расстояния между генами
дрозофилы
Slide 26
4 степени надежности
локализации данного гена
подтвержденная (установлена в двух и
более независимых лабораториях или на
материале двух и более независимых тестобъектов),
предварительная (1 лаборатория или 1
анализируемая семья),
противоречивая (несовпадение данных
разных исследователей),
сомнительная (не уточненные
окончательно данные одной лаборатории)
Slide 27
Минусы:
частота рекомбинации в
разных точках генома
различна, и расстояние
может существенно
варьировать
Необходим
тщательный
анализ
родословной
(если
картируется ген
заболевания)
в результате карты
сцеплений не отражают
реальных физических
расстояний между
маркерами и генами
на хромосомах.
Slide 28
Цитогенетическое
картирование
осуществляется с применением
методов цитогенетики, когда для
локализации каких-либо
нуклеотидных
последовательностей и
определения их взаимного
расположения используются
цитологические препараты
Slide 29
Цитологические карты
Метод цитологических карт основан на
использовании хромосомных перестроек –
перекрывающихся делеций.
При облучении и действии других
мутагенов в хромосомах часто
наблюдаются потери (делеции)
или вставки (дупликации)
небольших фрагментов,
сравнимых по величине с одним
или несколькими локусами.
Slide 30
Принципы:
Используются гетерозиготы по хромосомам, одна из которых
будет нести группу следующих друг за другом доминантных
аллелей, а гомологичная ей - группу рецессивных аллелей тех же
генов.
Если в хромосоме с доминантными генами произошла утрата
отдельных генов, например DE, то у гетерозиготы ABC/abcde будут
проявляться рецессивные признаки de. На этом принципе основан
метод перекрывающихся делеции, используемый при построении
цитологических карт.
Slide 31
Методы
дифференциального
окрашивания позволяют
идентифицировать на
препарате как отдельную
хромосому, так и любой
участок хромосомы
Разработанный на дрозофиле
специальный метод
перекрывающихся делеций был
использован для
цитологического картирования
генов у представителей многих
видов.
Slide 32
Хромосомные комплексы китайского хомячка
(А), мыши (Б) и их соматического гибрида (В)
Slide 33
Сравнение генетических и
цитологических карт хромосом
показывает их соответствие:
чем больший процент
кроссинговера разделяет пару
генов, тем больше и физическое
расстояние между ними.
Slide 34
Запись локализации
гена
Согласно официально утвержденной номенклатуре
(ISCN,1978), каждая хромосома человека после
дифференциальной окраски может быть разделена на
, нумерация которых начинается от
центромеры вверх (
), либо вниз
).
в каждом
участке тоже нумеруются в аналогичном порядке. Крупные
полосы разделяются на более мелкие
Slide 35
Slide 36
Алгоритм решения
задач на картирование
генов
Slide 37
Пример:
Составьте карту хромосомы,
содержащую гены, если
частота кроссинговера между
генами и равна 2,5%, и -
3,7%, и -6%, и - 2,8%, и -
6,2%, и - 15%, и - 8,8%
Slide 38
Slide 39
Используемая
литература
Э. Р. Рахманалиев, Е. А. Климов, Г. Е. Сулимова МЕТОДЫ
КАРТИРОВАНИЯ ГЕНОМОВ МЛЕКОПИТАЮЩИХ.
КАРТИРОВАНИЕ С ИСПОЛЬЗОВАНИЕМ РАДИАЦИОННЫХ
ГИБРИДОВ (RH КАРТИРОВАНИЕ)
Аксенович Т.И. Проблемы картирования QTL (Институт
цитологии и генетики СО РАН, Новосибирск)
Мяндлина Г.И. Молекулярные основы медицинской
генетики(кафедра биологии и общей генетики,
медицинского факультета РУДН)
В.И. Иванов Генетика Учебник для вузов, 2006
 Генетические и физические карты По общепринятой классификации методы картирования геномов подразделяют на две категории: p Генетическое картирование p Физическое картирование 2
Генетические и физические карты По общепринятой классификации методы картирования геномов подразделяют на две категории: p Генетическое картирование p Физическое картирование 2
 Составление генетических карт p p Маркёры – позиции каких-либо отличительных признаков. В качестве маркёров на протяжении десятилетий использовались гены, определяющие легко различимые фенотипы. Для более сложных карт использовались в качестве фенотипических признаков организма его биохимические особенности. Карта, основанная на генах, не может быть очень подробной. Также только часть всего числа генов существует в удобно различимых аллельных формах. 3
Составление генетических карт p p Маркёры – позиции каких-либо отличительных признаков. В качестве маркёров на протяжении десятилетий использовались гены, определяющие легко различимые фенотипы. Для более сложных карт использовались в качестве фенотипических признаков организма его биохимические особенности. Карта, основанная на генах, не может быть очень подробной. Также только часть всего числа генов существует в удобно различимых аллельных формах. 3
 ДНК-маркёры ДНК–маркёры – нанесённые на карту особенности, которые не являются генами. Всякий пригодный ДНК–маркёр должен иметь два аллеля, как и ген–маркёр. Этому требованию удовлетворяют три типа особенностей последовательности ДНК: -Полиморфизмы длины фрагментов рестрикции (RFLP); -Полиморфизмы длины простых последовательностей (SSLP); -Полиморфизмы отдельных нуклеотидов (SNP). 4
ДНК-маркёры ДНК–маркёры – нанесённые на карту особенности, которые не являются генами. Всякий пригодный ДНК–маркёр должен иметь два аллеля, как и ген–маркёр. Этому требованию удовлетворяют три типа особенностей последовательности ДНК: -Полиморфизмы длины фрагментов рестрикции (RFLP); -Полиморфизмы длины простых последовательностей (SSLP); -Полиморфизмы отдельных нуклеотидов (SNP). 4
 1 ДНК-маркёр. Полиморфизмы длины рестриктов RFLP – первый тип ДНК-маркёров, который был полностью изучен. Ферменты рестрикции разрезают ДНК в определённых сайтах узнавания. Эта специфичность означает, что обработка молекулы ДНК ферментом рестрикции должна производить один и тот же набор фрагментов. С молекулами геномной ДНК так происходит не всегда, так как некоторые участки рестрикции полиморфны и существуют в виде двух аллелей: один показывает правильную последовательность для участка рестрикции и потому разрезается ферментом при обработке ДНК, а второй аллель несёт видоизменение последовательности, так что участок рестрикции уже не опознаётся. В результате этого два смежных фрагмента 5 рестрикции остаются связанными вместе, что
1 ДНК-маркёр. Полиморфизмы длины рестриктов RFLP – первый тип ДНК-маркёров, который был полностью изучен. Ферменты рестрикции разрезают ДНК в определённых сайтах узнавания. Эта специфичность означает, что обработка молекулы ДНК ферментом рестрикции должна производить один и тот же набор фрагментов. С молекулами геномной ДНК так происходит не всегда, так как некоторые участки рестрикции полиморфны и существуют в виде двух аллелей: один показывает правильную последовательность для участка рестрикции и потому разрезается ферментом при обработке ДНК, а второй аллель несёт видоизменение последовательности, так что участок рестрикции уже не опознаётся. В результате этого два смежных фрагмента 5 рестрикции остаются связанными вместе, что


 Визуализация RFLP 2. ПЦР используется гораздо чаще. Затравки для ПЦР разрабатываются таким образом, чтобы они отжигались к обеим сторонам полиморфного участка, и RFLP типизируется путём обработки размноженного фрагмента ферментом рестрикции и последующего пробега образца в агарозном геле. 8
Визуализация RFLP 2. ПЦР используется гораздо чаще. Затравки для ПЦР разрабатываются таким образом, чтобы они отжигались к обеим сторонам полиморфного участка, и RFLP типизируется путём обработки размноженного фрагмента ферментом рестрикции и последующего пробега образца в агарозном геле. 8
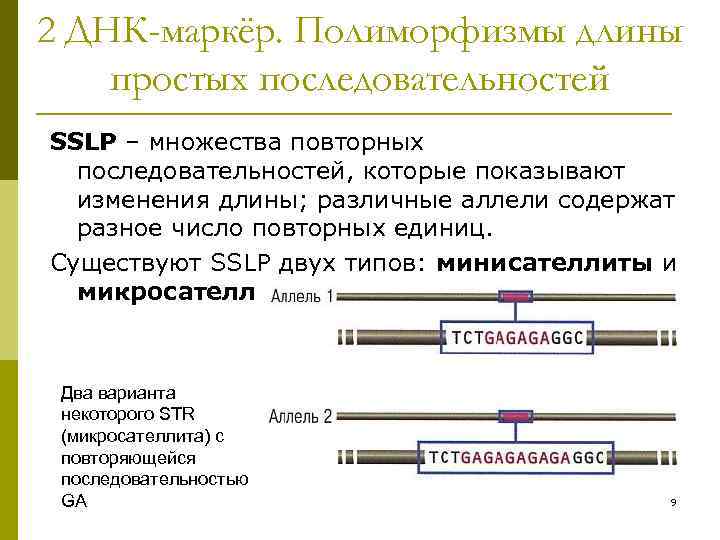 2 ДНК-маркёр. Полиморфизмы длины простых последовательностей SSLP – множества повторных последовательностей, которые показывают изменения длины; различные аллели содержат разное число повторных единиц. Существуют SSLP двух типов: минисателлиты и микросателлиты. Два варианта некоторого STR (микросателлита) с повторяющейся последовательностью GA 9
2 ДНК-маркёр. Полиморфизмы длины простых последовательностей SSLP – множества повторных последовательностей, которые показывают изменения длины; различные аллели содержат разное число повторных единиц. Существуют SSLP двух типов: минисателлиты и микросателлиты. Два варианта некоторого STR (микросателлита) с повторяющейся последовательностью GA 9
 Типы SSLP Минисателлиты (переменное число тандемных повторений, или VNTR). Повторная единица может иметь длину до 25 п. н. 2. Микросателлиты (простые тандемные повторения, или STR). Повторяющийся элемент – 13 п. н. или меньше. 1. 10
Типы SSLP Минисателлиты (переменное число тандемных повторений, или VNTR). Повторная единица может иметь длину до 25 п. н. 2. Микросателлиты (простые тандемные повторения, или STR). Повторяющийся элемент – 13 п. н. или меньше. 1. 10
 Типы SSLP ДНК-маркёры, основанные на микросателлитах, более популярны, чем основанные на минисателлитах, по двум причинам: -Минисателлиты неравномерно распределены по всему геному, чаще встречаются в теломерных областях на концах хромосом, микросателлиты более равномерно распределены в геноме. -точная типизация полиморфизма длины путём ПЦР возможна при длине последовательностей не более 300 п. н. , а большинство минисателлитных аллелей 11
Типы SSLP ДНК-маркёры, основанные на микросателлитах, более популярны, чем основанные на минисателлитах, по двум причинам: -Минисателлиты неравномерно распределены по всему геному, чаще встречаются в теломерных областях на концах хромосом, микросателлиты более равномерно распределены в геноме. -точная типизация полиморфизма длины путём ПЦР возможна при длине последовательностей не более 300 п. н. , а большинство минисателлитных аллелей 11
 3 ДНК- маркёр. Полиморфизмы отдельных нуклеотидов SNP – это позиции генома, в которых некоторые индивидуумы имеют один нуклеотид, например, G, а другие имеют отличающийся от него нуклеотид – С. 12
3 ДНК- маркёр. Полиморфизмы отдельных нуклеотидов SNP – это позиции генома, в которых некоторые индивидуумы имеют один нуклеотид, например, G, а другие имеют отличающийся от него нуклеотид – С. 12
 Большинство SNP имеют 2 аллеля, поскольку SNP возникают, когда в геноме происходят точечные мутации, преобразовывающие один нуклеотид в другой. Если подобная мутация случится в репродуктивных клетках, то один или более его потомков могут унаследовать эту мутацию, и в итоге SNP станет закреплённым в популяции. 13
Большинство SNP имеют 2 аллеля, поскольку SNP возникают, когда в геноме происходят точечные мутации, преобразовывающие один нуклеотид в другой. Если подобная мутация случится в репродуктивных клетках, то один или более его потомков могут унаследовать эту мутацию, и в итоге SNP станет закреплённым в популяции. 13
 Методы типизации SNP Методы базируются на анализе гибридизацией олигонуклеотидов. -Технология чипов ДНК - Методы гибридизации в растворе - Анализ лигированием олигонуклеотидов (OLA) - Система размножения термостабильных мутаций, или тест ARMS. 14
Методы типизации SNP Методы базируются на анализе гибридизацией олигонуклеотидов. -Технология чипов ДНК - Методы гибридизации в растворе - Анализ лигированием олигонуклеотидов (OLA) - Система размножения термостабильных мутаций, или тест ARMS. 14
 Методы типизации SNP Технология чипов ДНК На поверхность стеклянной пластины площадью 2 см 2 с множеством различных олигонуклеотидов пипеткой наносится предназначенная для тестирования ДНК, помеченная флюоресцентным маркёром. Гибридизация детектируется путём анализа чипа с помощью флюоресцентного микроскопа. Позиции, в которых испускается флюоресцентный сигнал, показывают, какие олигонуклеотиды 1. 15
Методы типизации SNP Технология чипов ДНК На поверхность стеклянной пластины площадью 2 см 2 с множеством различных олигонуклеотидов пипеткой наносится предназначенная для тестирования ДНК, помеченная флюоресцентным маркёром. Гибридизация детектируется путём анализа чипа с помощью флюоресцентного микроскопа. Позиции, в которых испускается флюоресцентный сигнал, показывают, какие олигонуклеотиды 1. 15
 Методы типизации SNP 2. Метод гибридизации в растворе Используется пара меток, в которые входит флюоресцентный краситель и вещество, гасящее флюоресцентный сигнал при сближении с испускающим его красителем. Краситель прикрепляется к одному концу олигонуклеотида, гасящее вещество – к другому концу. Если между олигонуклеотидом и тестируемой ДНК происходит гибридизация, то данное спаривание оснований нарушается, гаситель отрывается от красителя, и тот вырабатывает флюоресцентный сигнал. 16
Методы типизации SNP 2. Метод гибридизации в растворе Используется пара меток, в которые входит флюоресцентный краситель и вещество, гасящее флюоресцентный сигнал при сближении с испускающим его красителем. Краситель прикрепляется к одному концу олигонуклеотида, гасящее вещество – к другому концу. Если между олигонуклеотидом и тестируемой ДНК происходит гибридизация, то данное спаривание оснований нарушается, гаситель отрывается от красителя, и тот вырабатывает флюоресцентный сигнал. 16
 Методы типизации SNP 3. Анализ лигированием нуклеотидов (OLA) Применяются два олигонуклеотида, которые отжигаются смежно другу, при этом 3’-конец одного из них точно попадает в SNP. Этот олигонуклеотид образует полностью спаренную основаниями структуру, если в матричной ДНК присутствует одна версия SNP, и когда это происходит, данный олигонуклеотид может быть 17
Методы типизации SNP 3. Анализ лигированием нуклеотидов (OLA) Применяются два олигонуклеотида, которые отжигаются смежно другу, при этом 3’-конец одного из них точно попадает в SNP. Этот олигонуклеотид образует полностью спаренную основаниями структуру, если в матричной ДНК присутствует одна версия SNP, и когда это происходит, данный олигонуклеотид может быть 17
 Методы типизации SNP 4. Система размножения термостабильных мутаций (тест ARMS) Контрольным олигонуклеотидом выступает одна из пары затравок ПЦР. Если контрольная затравка отжигается к SNP, то он может быть продолжен с помощью полимеразы Taq и ПЦР может иметь место, но если она не отжигается из-за того, что присутствует альтернативная версия SNP, то никаких продуктов ПЦР не 18
Методы типизации SNP 4. Система размножения термостабильных мутаций (тест ARMS) Контрольным олигонуклеотидом выступает одна из пары затравок ПЦР. Если контрольная затравка отжигается к SNP, то он может быть продолжен с помощью полимеразы Taq и ПЦР может иметь место, но если она не отжигается из-за того, что присутствует альтернативная версия SNP, то никаких продуктов ПЦР не 18
 Сцепление генетических признаков Генетическое картирование основано на законах наследственности, описанных Грегором Менделем ещё в 1865 году. Помимо первых двух законов Менделя, встречаются ещё два случая необычного сцепления: -Неполное доминирование (гетерозиготная форма проявляет фенотип, промежуточный между двумя гомозиготными формами); -Кодоминирование (гетерозиготная форма показывает оба гомозиготных фенотипа) 19
Сцепление генетических признаков Генетическое картирование основано на законах наследственности, описанных Грегором Менделем ещё в 1865 году. Помимо первых двух законов Менделя, встречаются ещё два случая необычного сцепления: -Неполное доминирование (гетерозиготная форма проявляет фенотип, промежуточный между двумя гомозиготными формами); -Кодоминирование (гетерозиготная форма показывает оба гомозиготных фенотипа) 19
 Определяющий шаг в развитии генетического картирования Когда в 1900 году законы Менделя были переоткрыты, выяснилось, что полное сцепление, которое ожидалось между многими парами генов, не осуществилось. Пары генов или наследовались независимо, или показывали лишь неполное сцепление: иногда наследовались вместе, иногда порознь. p Разрешение этого противоречия и стало решающим шагом в развитии составления генетических карт. p 20
Определяющий шаг в развитии генетического картирования Когда в 1900 году законы Менделя были переоткрыты, выяснилось, что полное сцепление, которое ожидалось между многими парами генов, не осуществилось. Пары генов или наследовались независимо, или показывали лишь неполное сцепление: иногда наследовались вместе, иногда порознь. p Разрешение этого противоречия и стало решающим шагом в развитии составления генетических карт. p 20
 Рассуждения Томаса Моргана Неполное сцепление объясняется поведением хромосом во время мейоза. p Процесс кроссинговера (или рекомбинации) был открыт бельгийским цитологом Янсеном в 1909 году и помог Моргану объяснить неполное сцепление. Рассмотрим эффект, который имеет кроссинговер на наследование генов. p 21
Рассуждения Томаса Моргана Неполное сцепление объясняется поведением хромосом во время мейоза. p Процесс кроссинговера (или рекомбинации) был открыт бельгийским цитологом Янсеном в 1909 году и помог Моргану объяснить неполное сцепление. Рассмотрим эффект, который имеет кроссинговер на наследование генов. p 21
 Эффект кроссинговера Имеется два возможных сценария: p Между генами А и В не происходит кроссинговер. Тогда две гаметы имеют генотип АВ, две другие – аb. p Между генами А и В происходит кроссинговер. Это приводит к обмену сегментами ДНК между гомологичными хромосомами. В итоге каждая гамета имеет отличный от других генотип: AB, a. B, Ab и ab. Помимо гамет с родительскими генотипами появляются гаметы с 22
Эффект кроссинговера Имеется два возможных сценария: p Между генами А и В не происходит кроссинговер. Тогда две гаметы имеют генотип АВ, две другие – аb. p Между генами А и В происходит кроссинговер. Это приводит к обмену сегментами ДНК между гомологичными хромосомами. В итоге каждая гамета имеет отличный от других генотип: AB, a. B, Ab и ab. Помимо гамет с родительскими генотипами появляются гаметы с 22
 Составление генетических карт Когда Морган объяснил неполное сцепление кроссинговером, он изобрёл способ наносить на карту отдельные позиции генов в хромосоме. Допустим, кроссинговер является случайным событием, а значит, может произойти в любой позиции на протяжении пары вытянутых одна вдоль другой хроматид. Если это верно, то два гена, расположенные близко друг к другу, будут разделяться кроссинговерами реже, чем гены, лежащие дальше друг от друга. Частота, с которой гены разъединяются кроссинговерами, будет прямо пропорциональна отдалению их друг от друга. Поэтому частота рекомбинации является мерой расстояния 23
Составление генетических карт Когда Морган объяснил неполное сцепление кроссинговером, он изобрёл способ наносить на карту отдельные позиции генов в хромосоме. Допустим, кроссинговер является случайным событием, а значит, может произойти в любой позиции на протяжении пары вытянутых одна вдоль другой хроматид. Если это верно, то два гена, расположенные близко друг к другу, будут разделяться кроссинговерами реже, чем гены, лежащие дальше друг от друга. Частота, с которой гены разъединяются кроссинговерами, будет прямо пропорциональна отдалению их друг от друга. Поэтому частота рекомбинации является мерой расстояния 23
 Анализ сцепления генетических признаков у организмов различного типа. Включает три ситуации: p Анализ сцепления генетических признаков у видов наподобие плодовой мушки и мыши, с которыми можно выполнять эксперименты по скрещиванию; p Анализ сцепления генетических признаков у людей, с которыми нельзя проводить эксперименты, но можно изучать родословные; p Анализ сцепления генетических признаков у бактерий, которые не 24
Анализ сцепления генетических признаков у организмов различного типа. Включает три ситуации: p Анализ сцепления генетических признаков у видов наподобие плодовой мушки и мыши, с которыми можно выполнять эксперименты по скрещиванию; p Анализ сцепления генетических признаков у людей, с которыми нельзя проводить эксперименты, но можно изучать родословные; p Анализ сцепления генетических признаков у бактерий, которые не 24
 Анализ сцепления генетических признаков при возможности проведения скрещивания Метод основан на анализе потомства от экспериментальных скрещиваний, при известных генотипах родителей. Обычно используется анализирующее скрещивание. Этот метод применим ко всем эукариотам, но неприменим к человеку из этических соображений. 25
Анализ сцепления генетических признаков при возможности проведения скрещивания Метод основан на анализе потомства от экспериментальных скрещиваний, при известных генотипах родителей. Обычно используется анализирующее скрещивание. Этот метод применим ко всем эукариотам, но неприменим к человеку из этических соображений. 25
 Составление генетической карты на основе анализа родословной человека Зачастую из-за соблюдения научной и медицинской этики учёные могут оперировать лишь скудными данными, так как браки редко дают удобное анализирующее скрещивание, и генотипы многих членов семей могут быть неизвестны ввиду смерти или нежелания сотрудничать. Обыкновенно, чтобы решить необходимую генетическую задачу, достаточно знать дополнительно генотип хотя бы одного родственника, но по разным причинам это невозможно. 26
Составление генетической карты на основе анализа родословной человека Зачастую из-за соблюдения научной и медицинской этики учёные могут оперировать лишь скудными данными, так как браки редко дают удобное анализирующее скрещивание, и генотипы многих членов семей могут быть неизвестны ввиду смерти или нежелания сотрудничать. Обыкновенно, чтобы решить необходимую генетическую задачу, достаточно знать дополнительно генотип хотя бы одного родственника, но по разным причинам это невозможно. 26
 Составление генетических карт бактерий Главная трудность состоит в том, что бактерии гаплоидны и не подвергаются мейозу. Поэтому используются три способа, способные вызвать кроссинговер: p В процессе коньюгации происходит передача эписомы (сегмент хромосомной ДНК длиной до 1 млн. п. н.) p Трансдукция (передача фрагмента ДНК длиной до 50 тыс. п. н. через бактериофаг) p Трансформация (клетка-реципиент 27
Составление генетических карт бактерий Главная трудность состоит в том, что бактерии гаплоидны и не подвергаются мейозу. Поэтому используются три способа, способные вызвать кроссинговер: p В процессе коньюгации происходит передача эписомы (сегмент хромосомной ДНК длиной до 1 млн. п. н.) p Трансдукция (передача фрагмента ДНК длиной до 50 тыс. п. н. через бактериофаг) p Трансформация (клетка-реципиент 27
 Составление физических карт Полученная исключительно генетическими методами карта не будет полностью точна. Это обусловлено следующими причинами: 1. Разрешение генетической карты зависит от числа кроссинговеров, которые были набраны. Для микроорганизмов это не главная проблема, поскольку они могут быть получены в любом количестве. Проблема с людьми и другими эукариотами в том, что невозможно получить большое число потомков, так как может быть изучено сравнительно 28
Составление физических карт Полученная исключительно генетическими методами карта не будет полностью точна. Это обусловлено следующими причинами: 1. Разрешение генетической карты зависит от числа кроссинговеров, которые были набраны. Для микроорганизмов это не главная проблема, поскольку они могут быть получены в любом количестве. Проблема с людьми и другими эукариотами в том, что невозможно получить большое число потомков, так как может быть изучено сравнительно 28
 Составление физических карт 2. Генетические карты имеют ограниченную точность. На картинке изображено сравнение физической и генетической карт дрожжей Saccharomyces cerevisiae. Сравнение показывает, что порядок двух верхних маркёров на генетической карте неверен и также есть различия в относительном 29
Составление физических карт 2. Генетические карты имеют ограниченную точность. На картинке изображено сравнение физической и генетической карт дрожжей Saccharomyces cerevisiae. Сравнение показывает, что порядок двух верхних маркёров на генетической карте неверен и также есть различия в относительном 29

 Составление рестрикционных карт Простейший способ составления рестрикционной карты – сравнение размеров фрагментов, полученных при переваривании молекулы ДНК двумя разными ферментами рестрикции. Выбрать единственно верную карту позволяет дополнительная обработка исходной ДНК одним ферментом с предотвращением протекания переваривания до конца. Это называется частичной рестрикцией. Масштаб рестрикционной карты ограничивается длиной рестриктов. Рестрикционное картирование более приемлемо для маленьких молекул. 31
Составление рестрикционных карт Простейший способ составления рестрикционной карты – сравнение размеров фрагментов, полученных при переваривании молекулы ДНК двумя разными ферментами рестрикции. Выбрать единственно верную карту позволяет дополнительная обработка исходной ДНК одним ферментом с предотвращением протекания переваривания до конца. Это называется частичной рестрикцией. Масштаб рестрикционной карты ограничивается длиной рестриктов. Рестрикционное картирование более приемлемо для маленьких молекул. 31
 Составление рестрикционных карт Возможно ли использование рестрикционного анализа для картирования геномов размером более 50 тыс. п. н. ? Да, ограничения рестрикционного картирования могут быть ослаблены за счёт подбора ферментов, которые имеют редкие участки разрезания в целевой молекуле ДНК («редкощепящие рестриктазы») 32
Составление рестрикционных карт Возможно ли использование рестрикционного анализа для картирования геномов размером более 50 тыс. п. н. ? Да, ограничения рестрикционного картирования могут быть ослаблены за счёт подбора ферментов, которые имеют редкие участки разрезания в целевой молекуле ДНК («редкощепящие рестриктазы») 32
 Метод OFAGE Электрофорез в геле с ортогонально чередующимся полем. Таким образом, каждое изменение поля вынуждает молекулы перестраиваться, короткие молекулы перестраиваются и Электрическое поле чередуется мигрируют через гель между двумя парами быстрее длинных. За счёт электродов, каждая из которых такого приёма позволяются помещена по углом 45° к более длинные фрагменты, продольной линии геля. чем при обычном электрофорезе. К подобного рода методам относят также CHEF – электрофорез в геле с однородными электрическими полями и 33 FIGE – электрофорез в геле с обращением поля.
Метод OFAGE Электрофорез в геле с ортогонально чередующимся полем. Таким образом, каждое изменение поля вынуждает молекулы перестраиваться, короткие молекулы перестраиваются и Электрическое поле чередуется мигрируют через гель между двумя парами быстрее длинных. За счёт электродов, каждая из которых такого приёма позволяются помещена по углом 45° к более длинные фрагменты, продольной линии геля. чем при обычном электрофорезе. К подобного рода методам относят также CHEF – электрофорез в геле с однородными электрическими полями и 33 FIGE – электрофорез в геле с обращением поля.
 Непосредственное наблюдение участков рестрикции в молекулах ДНК. Для нанесения на карту участков рестрикции можно использовать методы, не связанные с электрофорезом. p Метод оптического картирования: позиции участков рестрикции определяются путём непосредственного наблюдения разрезанных молекул ДНК в микроскоп. Для закрепления ДНК на предметном стекле используют вытягивание гелем и расчёсывание молекул. p Для вытягивания гелем хромосомную ДНК переводят во взвесь в расплавленной агарозе и помещают на предметное стекло микроскопа. По мере охлаждения и затвердевания геля молекулы ДНК вытягиваются. p Для расчёсывания волокна ДНК приготовляют путём погружения покрытого силиконом покровного стекла в раствор ДНК, выдерживая его там в течение 5 минут. Далее вынимают стекло из раствора. Сила, необходимая для 34 протягивания ДНК через поверхностный мениск, заставляет каждую из них вытягиваться в линию. При засыхании ДНК
Непосредственное наблюдение участков рестрикции в молекулах ДНК. Для нанесения на карту участков рестрикции можно использовать методы, не связанные с электрофорезом. p Метод оптического картирования: позиции участков рестрикции определяются путём непосредственного наблюдения разрезанных молекул ДНК в микроскоп. Для закрепления ДНК на предметном стекле используют вытягивание гелем и расчёсывание молекул. p Для вытягивания гелем хромосомную ДНК переводят во взвесь в расплавленной агарозе и помещают на предметное стекло микроскопа. По мере охлаждения и затвердевания геля молекулы ДНК вытягиваются. p Для расчёсывания волокна ДНК приготовляют путём погружения покрытого силиконом покровного стекла в раствор ДНК, выдерживая его там в течение 5 минут. Далее вынимают стекло из раствора. Сила, необходимая для 34 протягивания ДНК через поверхностный мениск, заставляет каждую из них вытягиваться в линию. При засыхании ДНК

 Флюоресцентная гибридизация in situ (FISH) В этой методике маркёром является последовательность ДНК, которая отображается путём гибридизации с флюоресцентным зондом. Ненарушенная хромосома исследуется путём её зондирования меченой молекулой ДНК. Для работы метода ДНК в хромосоме денатурируется(высушивается на предметном стекле и обрабатывается 36
Флюоресцентная гибридизация in situ (FISH) В этой методике маркёром является последовательность ДНК, которая отображается путём гибридизации с флюоресцентным зондом. Ненарушенная хромосома исследуется путём её зондирования меченой молекулой ДНК. Для работы метода ДНК в хромосоме денатурируется(высушивается на предметном стекле и обрабатывается 36
 FISH в действии 1. 2. Первоначально метод использовался с метафазными хромосомами, но их сильная уплотнённость не позволяла составлять карты с высокой разрешающей способностью. В 1995 году был разработан ряд методов FISH более высокого разрешения. Оно достигалось за счёт изменения характера изучаемого хромосомного аппарата. Если метафазные хромосомы слишком сжаты для крупномасштабного картирования, то нам следует использовать хромосомы в более вытянутом виде. Добиться этого можно двумя способами. Механически вытянутые хромосомы могут быть получены за счёт изменения метода приготовления препарата, применяемого для выделения хромосом из метафазных ядер. Неметафазные хромосомы используются, потому что во всех стадиях клеточного цикла, кроме метафазы, хромосомы пребывают в естественном для них 37 развёрнутом состоянии. Интерфазные хромосомы
FISH в действии 1. 2. Первоначально метод использовался с метафазными хромосомами, но их сильная уплотнённость не позволяла составлять карты с высокой разрешающей способностью. В 1995 году был разработан ряд методов FISH более высокого разрешения. Оно достигалось за счёт изменения характера изучаемого хромосомного аппарата. Если метафазные хромосомы слишком сжаты для крупномасштабного картирования, то нам следует использовать хромосомы в более вытянутом виде. Добиться этого можно двумя способами. Механически вытянутые хромосомы могут быть получены за счёт изменения метода приготовления препарата, применяемого для выделения хромосом из метафазных ядер. Неметафазные хромосомы используются, потому что во всех стадиях клеточного цикла, кроме метафазы, хромосомы пребывают в естественном для них 37 развёрнутом состоянии. Интерфазные хромосомы
 Картирование с помощью меченых участков последовательности (STS) В настоящее время это самый мощный метод физического картирования. Меченый участок последовательности, или STS – короткая последовательность ДНК, 100 -500 п. н. в длину, которая легко опознаётся и лишь единожды встречается в хромосоме или изучаемом геноме. Чтобы нанести на карту набор STS, необходимо располагать множеством перекрывающихся фрагментов ДНК из отдельной хромосомы или полного генома. Какие фрагменты содержат какие STS, определяется методом гибридизационного анализа, или, чаще, ПЦР. Любая уникальная последовательность ДНК может быть использована в качестве STS. Для этого последовательность ДНК должна быть известна, а STS должен иметь уникальное 38 местоположение на изучаемой хромосоме.
Картирование с помощью меченых участков последовательности (STS) В настоящее время это самый мощный метод физического картирования. Меченый участок последовательности, или STS – короткая последовательность ДНК, 100 -500 п. н. в длину, которая легко опознаётся и лишь единожды встречается в хромосоме или изучаемом геноме. Чтобы нанести на карту набор STS, необходимо располагать множеством перекрывающихся фрагментов ДНК из отдельной хромосомы или полного генома. Какие фрагменты содержат какие STS, определяется методом гибридизационного анализа, или, чаще, ПЦР. Любая уникальная последовательность ДНК может быть использована в качестве STS. Для этого последовательность ДНК должна быть известна, а STS должен иметь уникальное 38 местоположение на изучаемой хромосоме.
 Методы получения STS 1. 2. 3. Ярлыки экспрессируемых последовательностей – короткие последовательности, получаемые анализом клонов к. ДНК. Полиморфизмы длины простой последовательности (SSLP) Случайные геномные последовательности – получают секвенированием случайных частей клонированной геномной к. ДНК. 39
Методы получения STS 1. 2. 3. Ярлыки экспрессируемых последовательностей – короткие последовательности, получаемые анализом клонов к. ДНК. Полиморфизмы длины простой последовательности (SSLP) Случайные геномные последовательности – получают секвенированием случайных частей клонированной геномной к. ДНК. 39
 Фрагменты ДНК для картирования с помощью STS Иначе реактив для картирования; существуют в виде библиотеки клонов и радиационных гибридов. Радиационный гибрид – клетка грызуна, содержащая фрагменты хромосом другого организма. При разбиении хромосомы на фрагменты большая доза излучения давала большее число фрагментов. Слияние стимулируется химически (полиэтиленгликолем) или биологически – вирусом Сёндай. 40
Фрагменты ДНК для картирования с помощью STS Иначе реактив для картирования; существуют в виде библиотеки клонов и радиационных гибридов. Радиационный гибрид – клетка грызуна, содержащая фрагменты хромосом другого организма. При разбиении хромосомы на фрагменты большая доза излучения давала большее число фрагментов. Слияние стимулируется химически (полиэтиленгликолем) или биологически – вирусом Сёндай. 40
 Выводы p p p Карты геномов – опорная схема для проектов секвенирования, так как они позволяют проверять точность собранной последовательности ДНК. Генетические карты строят по результатам экспериментов по скрещиванию и анализа родословных, физические карты – посредством прямого наблюдения молекул ДНК. В самых первых генетических картах маркёрами выступали гены, аллели которых можно было легко отличать (по резко отличным фенотипам), ныне же ДНК-маркёрами являются полиморфизмы длины фрагмента рестрикции (RFLP), полиморфизмы длины простой последовательности (SSLP) и полиморфизмы отдельных нуклеотидов (SNP). Все они легко типизируются посредством ПЦР. Анализ сцепления генетических признаков позволяет определить частоту рекомбинации между парой маркёров. Для многих организмов анализ сцепления генетических признаков прослеживается при помощи запланированных 41 экспериментов по скрещиванию. С людьми их проведение
Выводы p p p Карты геномов – опорная схема для проектов секвенирования, так как они позволяют проверять точность собранной последовательности ДНК. Генетические карты строят по результатам экспериментов по скрещиванию и анализа родословных, физические карты – посредством прямого наблюдения молекул ДНК. В самых первых генетических картах маркёрами выступали гены, аллели которых можно было легко отличать (по резко отличным фенотипам), ныне же ДНК-маркёрами являются полиморфизмы длины фрагмента рестрикции (RFLP), полиморфизмы длины простой последовательности (SSLP) и полиморфизмы отдельных нуклеотидов (SNP). Все они легко типизируются посредством ПЦР. Анализ сцепления генетических признаков позволяет определить частоту рекомбинации между парой маркёров. Для многих организмов анализ сцепления генетических признаков прослеживается при помощи запланированных 41 экспериментов по скрещиванию. С людьми их проведение
 Выводы p p p Генетическое картирование генома человека опирается на сведения, почёрпнутые из анализа родословной. Низкое разрешение генетических карт уточняется физическим картированием. В молекуле ДНК позиции участков рестрикции определяются рестрикционным картированием. Флюоресцентная гибридизация более продуктивная, в ней препарат зондируется маркёром, меченным флюоресцентной меткой. Позиция гибридизации определяется микроскопированием. Наиболее подробные физические карты получаются методом картирования содержания меченых участков последовательности (STS). Позиция маркёра на карте определяется фрагментами из коллекции, содержащими копии маркёра. 42
Выводы p p p Генетическое картирование генома человека опирается на сведения, почёрпнутые из анализа родословной. Низкое разрешение генетических карт уточняется физическим картированием. В молекуле ДНК позиции участков рестрикции определяются рестрикционным картированием. Флюоресцентная гибридизация более продуктивная, в ней препарат зондируется маркёром, меченным флюоресцентной меткой. Позиция гибридизации определяется микроскопированием. Наиболее подробные физические карты получаются методом картирования содержания меченых участков последовательности (STS). Позиция маркёра на карте определяется фрагментами из коллекции, содержащими копии маркёра. 42
Вскоре после переоткрытия законов Менделя немецкий цитолог Теодор Бовери (1902) представил доказательства в пользу участия хромосом в процессах наследственной передачи, показав, что нормальное развитие морского ежа возможно только при наличии всех хромосом. В это же время (1903 г.) американский цитолог Уильям Сэттон обратил внимание на параллелизм в поведении хромосом в мейозе и гипотетических факторов наследственности, существование которых предсказал еще сам Мендель.
Уильям Сэттон предположил, что в одной хромосоме может находиться несколько генов. В этом случае должно наблюдаться сцепленное наследование признаков, т.е. несколько разных признаков могут наследоваться так, как будто они контролируются одним геном. В 1906 г. У. Бэтсон и Р. Пеннет обнаружили сцепленное наследование у душистого горошка. Они изучали совместное наследование: окраски цветков (пурпурная или красная) и формы пыльцевых зерен (удлиненная или округлая). При скрещивании дигетерозигот в их потомстве наблюдалось расщепление 11,1:0,9:0,9:3,1 вместо ожидаемого 9:3:3:1. Создавалось впечатление, что факторы окраски и формы пыльцы имеют тенденцию при рекомбинации задатков оставаться вместе. Это явление авторы назвали «взаимным притяжением факторов», но природу его им выяснить не удалось.
Дальнейшее изучение хромосом как носителей информации происходило в первые десятилетия ХХ века в лаборатории Томаса Ханта Моргана (США) и его сотрудников (А. Стёртеванта, К. Бриджеса, Г. Мёллера). В качестве основного объекта исследований Морган использовал плодовую мушку дрозофилу (Drosophila melanogaster), которая оказалась очень удобным модельным объектом:
– Во-первых, эта мушка легко культивируется в лабораторных условиях.
– Во-вторых, она характеризуется малым числом хромосом 2 n = 8).
– В-третьих, в слюнных железах личинок дрозофилы имеются гигантские (политенные) хромосомы, удобные для прямого наблюдения.
– И, наконец, дрозофила отличается высокой изменчивостью морфологических признаков.
На основании экспериментов с плодовой мушкой дрозофилой Морганом и его учениками была разработана хромосомная теория наследственности.
Основные положения хромосомной теории наследственности:
1. Ген – это элементарный наследственный фактор (термин «элементарный» означает «неделимый без потери качества»). Ген представляет собой участок хромосомы, отвечающий за развитие определенного признака. Иначе говоря, гены локализованы в хромосомах.
2. В одной хромосоме могут содержаться тысячи генов, расположенных линейно (подобно бусинкам на нитке). Эти гены образуют группы сцепления. Число групп сцепления равно числу хромосом в гаплоидном наборе. Совокупность аллелей в одной хромосоме называется гаплотип. Примеры гаплотипов: ABCD (только доминантные аллели), abcd (только рецессивные аллели), AbCd (различные комбинации доминантных и рецессивных аллелей).
3. Если гены сцеплены между собой, то возникает эффект и сцепленного наследования признаков, т.е. несколько признаков наследуются так, как будто они контролируются одним геном. При сцепленном наследовании в череде поколений сохраняются исходные сочетания признаков.
4. Сцепление генов не абсолютно: в большинстве случаев гомологичные хромосомы обмениваются аллелями в результате перекреста (кроссинговера) в профазе первого деления мейоза. В результате кроссинговера образуются кроссоверные хромосомы (возникают новые гаплотипы, т.е. новые сочетания аллелей.). С участием кроссоверных хромосом в последующих поколениях у кроссоверных особей должны появляться новые сочетания признаков.
5. Вероятность появления новых сочетаний признаков вследствие кроссинговера прямо пропорциональна физическому расстоянию между генами. Это позволяет определять относительное расстояние между генами и строить генетические (кроссоверные) карты разных видов организмов.
КРОССИНГОВЕР
Кроссинговер (от англ. crossing-over – перекрёст) – это процесс обмена гомологичными участками гомологичных хромосом (хроматид).
Обычно кроссинговер происходит в мейозе I.
При кроссинговере происходит обмен генетическим материалом (аллелями) между хромосомами, и тогда происходит рекомбинация – появление новых сочетаний аллелей, например, AB + ab → Ab + aB.
Механизм кроссинговера «разрыв–воссоединение»
Согласно теории Янссенса–Дарлингтона, кроссинговер происходит в профазе мейоза. Гомологичные хромосомы с хроматидами АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные хроматиды Ab и аВ. В анафазе первого деления мейоза происходит расхождение двухроматидных хромосом, а во втором делении – расхождение хроматид (однохроматидных хромосом). Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи. Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи. Таким образом, вследствие кроссинговера происходит рекомбинация – появление новых сочетаний наследственных задатков в хромосомах.
Согласно другим теориям, кроссинговер связан с репликацией ДНК: или в пахитене мейоза, или в интерфазе. В частности, возможна смена матрицы в вилке репликации.
Генетические (кроссоверные) карты
Алфред Стёртевант (сотрудник Моргана) предположил, что частота кроссинговера на участке между генами, локализованными в одной хромосоме, может служить мерой расстояния между генами. Иными словами, частота кроссинговера, выражаемая отношением числа кроссоверных особей к общему числу особей, прямо пропорциональна расстоянию между генами. Тогда можно использовать частоту кроссинговера для того, чтобы определять взаимное расположение генов и расстояние между генами. Единицей расстояния между генами служит 1 % кроссинговера; в честь Моргана эта единица называется морганидой (М).
На основании генетического картирования составляются генетические карты – схемы, отражающие положение генов в хромосомах относительно других генов. На генетических картах крайнему гену (т.е. наиболее удаленному от центромеры) соответствует нулевая (исходная) точка. Удаленность какого-либо гена от нулевой точки обозначается в морганидах.
Построение генетических карт различных организмов имеет большое значение в здравоохранении, селекции и экологии. При изучении признаков человека (и в частности, генетических заболеваний) важно знать, какой именно ген определяет рассматриваемый признак. Эти знания позволяют составлять прогнозы при медико-генетическом консультировании, при разработке методов лечения генетических заболевания, в т.ч. и для коррекции генома. Знание генетических карт культурных растений и домашних животных позволяет планировать селекционный процесс, что способствует получению надежных результатов в краткие сроки. Построение генетических карт дикорастущих растений и диких животных важно и сточки зрения экологии. В частности, исследователь получает возможность изучать не просто фенотипические признаки организмов, а конкретные, генетически обусловленные признаки.
Двойной и множественный кроссинговер
Морган предположил, что кроссинговер между двумя генами может происходить не только в одной, но и в двух и даже большем числе точек. Четное число перекрестов между двумя генами, в конечном счете, не приводит к их перемещению из одной гомологичной хромосомы в другую, поэтому число кроссинговеров и, следовательно, расстояние между этими генами, определенное в эксперименте, снижаются. Обычно это относится к достаточно далеко расположенным друг от друга генам. Естественно, что вероятность двойного перекреста всегда меньше вероятности одинарного. В принципе она будет равна произведению вероятности двух единичных актов рекомбинации. Например, если одиночный перекрест будет происходить с частотой 0,2, то двойной – с частотой 0,2 × 0,2 = 0,04. В дальнейшем, наряду с двойным кроссинговером, было открыто и явление множественного кроссинговера: гомологичные хроматиды могут обмениваться участками в трех, четырех и более точках.
Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке происшедшего обмена.
Рассмотрим пример, описанный в одной из ранних работ Моргана. Он исследовал частоту кроссинговера между генами w (white – белые глаза), у (yellow – желтое тело) и m (miniature – маленькие крылья), локализованными в Х-хромосоме D. melanogaster. Расстояние между генами w и у в процентах кроссинговера составило 1,3, а между генами у и m – 32,6. Если два акта кроссинговера наблюдаются случайно, то ожидаемая частота двойного кроссинговера должна быть равна произведению частот кроссинговера между генами у и w и генами w и m. Другими словами, частота двойных кроссинговеров будет 0,43%. В действительности в опыте был обнаружен лишь один двойной кроссинговер на 2205 мух, т. е. 0,045%. Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления фактически наблюдаемой частоты двойного кроссинговера на теоретически ожидаемую (при отсутствии интерференции) частоту. Он назвал этот показатель коэффициентом коинциденции, т. е. совпадения. Меллер показал, что в Х-хромосоме дрозофилы интерференция особенно велика на небольших расстояниях; с увеличением интервала между генами интенсивность ее уменьшается и на расстоянии около 40 морганид и более коэффициент коинциденции достигает 1 (максимального своего значения).
Цитологическое доказательство кроссинговера
Прямые цитологические свидетельства обмена частей хромосом во время кроссинговера были получены в начале 30-х годов у дрозофилы и кукурузы.
Рассмотрим опыт Штерна, проведенный на D. melanogaster. Обычно две гомологичные хромосомы морфологически неразличимы. Штерн исследовал Х-хромосомы, которые имели морфологические различия и, следовательно, были гомологичны не полностью. Однако гомология между этими хромосомами сохранялась на большей части их длины, что позволяло им нормально спариваться и сегрегировать в мейозе (то есть нормально распределяться по дочерним клеткам). Одна из Х-хромосом самки в результате транслокации, т. е. перемещения фрагмента Y-хромосомы, приобрела Г-образную форму. Вторая Х-хромосома была короче нормальной, так как часть ее была перенесена на IV хромосому. Были получены самки, гетерозиготные по указанным двум, морфологически различным, Х-хромосомам, а также гетерозиготные по двум генам, локализованным в Х-хромосоме: Bar (В) и carnation (cr). Ген Bar – это полудоминантный ген, влияющий на количество фасеток и, следовательно, форму глаза (мутанты с аллелем В имеют полосковидные глаза). Ген cr контролирует окраску глаз (аллель cr+ обусловливает нормальную окраску глаз, а аллель cr – окраску глаз цвета красной гвоздики). Г-образная Х-хромосома несла аллели дикого типа В+ и cr+, укороченная хромосома – мутантные аллели В и cr. Самки указанного генотипа скрещивались с самцами, имевшими морфологически нормальную Х-хромосому с аллелями cr и В+. В потомстве самок было два класса мух с некроссоверными хромосомами (crB / crB+ и cr+B+ / crB+) и два класса мух, фенотип которых соответствовал кроссоверам (crB+ / crB+ и cr+B / crB+). Цитологическое исследование показало, что у кроссоверных особей произошел обмен участками Х-хромосом, и, соответственно, изменилась их форма. Все четыре класса самок имели по одной нормальной, т. е. палочковидной, хромосоме, полученной от отца. Кроссоверные самки содержали в своем кариотипе преобразованные в результате кроссинговера Х-хромосомы – длинную палочковидную или двуплечую с короткими плечами. Эти опыты, так же как и одновременно полученные аналогичные результаты на кукурузе, подтвердили гипотезу Моргана и его сотрудников о том, что кроссинговер представляет собой обмен участками гомологичных хромосом и что гены действительно локализованы в хромосомах.
Соматический (митотический) кроссинговер.
В соматических клетках иногда происходят обмены между хроматидами гомологичных хромосом, в результате которых наблюдается комбинативная изменчивость, подобная той, которая регулярно генерируется мейозом. Нередко, особенно у дрозофилы и низших эукариот, гомологичные хромосомы синаптируют в митозе. Одна из аутосомно-рецессивных мутаций человека, в гомозиготном состоянии приводящая к тяжелому заболеванию, известному под названием синдром Блюма, сопровождается цитологической картиной, напоминающей синапс гомологов и даже образование хиазм.
Доказательство митотического кроссинговера было получено на дрозофиле при анализе изменчивости признаков, определяемых генами у (yellow – желтое тело) и sn (singed – опаленные щетинки), которые находятся в Х-хромосоме. Самка с генотипом y sn+ / y+sn гетерозиготна по генам у и sn, и поэтому в отсутствие митотического кроссинговера ее фенотип будет нормальным. Однако если кроссинговер произошел на стадии четырех хроматид между хроматидами разных гомологов (но не между сестринскими хроматидами), причем место обмена находится между геном sn и центромерой, то образуются клетки с генотипами y sn+ / y+ sn+ и y+ sn / y+ sn. В этом случае на сером теле мухи с нормальными щетинками появятся близнецовые мозаичные пятна, одно из которых будет желтого цвета с нормальными щетинками, а другое - серого цвета с опаленными щетинками. Для этого необходимо, чтобы после кроссинговера обе хромосомы (бывшие хроматиды каждого из гомологов) y+ sn отошли к одному полюсу клетки, а хромосомы y sn+ – к другому. Потомки дочерних клеток, размножившись на стадии куколки, и приведут к появлению мозаичных пятен. Таким образом, мозаичные пятна образуются тогда, когда рядом расположены две группы (точнее, два клона) клеток, фенотипически отличающиеся друг от друга и от клеток остальных тканей данной особи.
Неравный кроссинговер
Это явление было детально изучено на примере гена Bar (В – полосковидные глаза), локализованного в Х-хромосоме D. melanogaster. Неравный кроссинговер связан с дупликацией какого-либо участка в одном из гомологов и с утратой его в другом гомологе. Обнаружено, что ген В может присутствовать в виде тандемных, т. е. следующих друг за другом, повторов, состоящих из двух и даже трех копий. Цитологический анализ подтвердил предположение о том, что неравный кроссинговер может вести к тандемным дупликациям. В области, соответствующей локализации гена В, на препаратах политенных хромосом отмечено увеличение числа дисков, пропорциональное дозе гена. Предполагается, что в эволюции неравный кроссинговер стимулирует создание тандемных дупликаций различных последовательностей и использование их в качестве сырого генетического материала для формирования новых генов и новых регуляционных систем.
Регуляция кроссинговера
Кроссинговер – это сложный физиолого-биохимический процесс, который находится под генетическим контролем клетки и подвержен влиянию факторов внешней среды. Поэтому в реальном эксперименте о частоте кроссинговера можно говорить, имея в виду все те условия, в которых она была определена. Кроссинговер практически отсутствует между гетероморфными Х- и Y-хромосомами. Если бы он происходил, то хромосомный механизм определения пола постоянно разрушался бы. Блокирование кроссинговера между этими хромосомами связано не только с различием в их величине (оно наблюдается не всегда), но и обусловлено Y-специфичными нуклеотидными последовательностями. Обязательное условие синапса хромосом (или их участков) - гомология нуклеотидных последовательностей.
Для абсолютного большинства высших эукариот характерна примерно одинаковая частота кроссинговера как у гомогаметного, так и гетерогаметного полов. Однако есть виды, у которых Кроссинговер отсутствует у особей гетерогаметного пола, в то время как у особей гомогаметного пола он протекает нормально. Такая ситуация наблюдается у гетерогаметных самцов дрозофилы и самок шелкопряда. Существенно, что частота митотического кроссинговера у этих видов у самцов и самок практически одинакова, что указывает на различные элементы контроля отдельных этапов генетической рекомбинации в половых и соматических клетках. В гетерохроматических районах, в частности прицентромерных, частота кроссинговера снижена, и поэтому истинное расстояние между генами в этих участках может быть изменено.
Обнаружены гены, выполняющие функции запирателей кроссинговера, но есть также гены, повышающие его частоту. Они иногда могут индуцировать заметное число кроссоверов у самцов дрозофилы. В качестве запирателей кроссинговера могут выступать также хромосомные перестройки, в частности инверсии. Они нарушают нормальную конъюгацию хромосом в зиготене.
Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток. Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
Биологическое значение кроссинговера
Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых представляет собой как единый суперген, контролирующий несколько признаков. В то же время, в ходе кроссинговера возникают рекомбинации – т.е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
Эволюционное значение сцепленного наследования. В результате сцепления одна хромосома может содержать как благоприятные аллели (например, А), так и нейтральные или относительно неблагоприятные (например, N). Если некоторый гаплотип (например, AN) повышает приспособленность его носителей за счет наличия благоприятных аллелей A, то в популяции будут накапливаться как благоприятные аллели, так и сцепленные с ними нейтральные или относительно неблагоприятные N.
Пример. Гаплотип AN обладает преимуществом перед гаплотипом “дикого типа» (++) за счет наличия благоприятного аллеля А, и тогда аллель N будет накапливаться в популяции, если он селективно нейтральный или даже относительно неблагоприятный (но его отрицательное влияние на приспособленность компенсируется положительным влиянием аллеля А).
Эволюционное значение кроссинговера. В результате кроссинговера неблагоприятные аллели, первоначально сцепленные с благоприятными, могут переходить в другую хромосому. Тогда возникают новые гаплотипы, не содержащие неблагоприятных аллелей, и эти неблагоприятные аллели элиминируются из популяции.
Пример. Гаплотип Al оказывается неблагоприятным по сравнению с гаплотипом «дикого типа» (++) за счет наличия летального аллеля l. Поэтому аллель А (благоприятный, нейтральный ил несколько снижающий приспособленность) не может проявиться в фенотипе, поскольку данный гаплотип (Al) содержит летальный аллель l. В результате кроссинговера возникают рекомбинантные гаплотипы A+ и +l. Гаплотип +l элиминируется из популяции, а гаплотип A+ фиксируется (даже в том случае, если аллель А несколько снижает приспособленность его носителей).
ДОПОЛНЕНИЯ
Принципы генетического картирования
Алфред Стёртевант (сотрудник Моргана) предположил, что частота кроссинговера на участке между генами, локализованными в одной хромосоме, может служить мерой расстояния между генами. Иными словами, частота кроссинговера, выражаемая отношением числа кроссоверных особей к общему числу особей, прямо пропорциональная расстоянию между генами. Тогда можно использовать частоту кроссинговера для того, чтобы определять взаимное расположение генов и расстояние между генами.
Генетическое картирование – это определение положения какого-либо гена по отношению к двум (как минимум) другим генам. Постоянство процента кроссинговера между определенными генами позволяет локализовать их. Единицей расстояния между генами служит 1 % кроссинговера; в честь Моргана эта единица называется морганидой (М).
На первом этапе картирования необходимо определить принадлежность гена к группе сцепления. Чем больше генов известно у данного вида, тем точнее результаты картирования. Все гены разбивают на группы сцепления. Число групп сцепления соответствует гаплоидному набору хромосом. Например, у D. melanogaster 4 группы сцепления, у кукурузы – 10, у мыши – 20, у человека – 23 группы сцепления. Как правило, число генов в группах сцепления зависит от линейных размеров соответствующих хромосом. Так, у плодовой мушки имеется одна (IV) точечная (при анализе в световом микроскопе) хромосома. Соответственно число генов в ней во много раз меньше, чем в остальных, значительно превосходящих ее по длине. Следует также отметить, что в гетерохроматических районах хромосом генов нет или почти нет, поэтому протяженные области конститутивного гетерохроматина могут несколько изменить пропорциональность числа генов и длины хромосомы.
На основании генетического картирования составляются генетические карты. На генетических картах крайнему гену (т.е. наиболее удаленному от центромеры) соответствует нулевая (исходная) точка. Удаленность какого-либо гена от нулевой точки обозначается в морганидах.
Если хромосомы достаточно длинные, то удаление гена от нулевой точки может превышать 50 М – тогда возникает противоречие между отмеченными на карте расстояниями, превышающими 50%, и постулированным выше положением, согласно которому 50 % кроссоверов, полученных в эксперименте, фактически должны означать отсутствие сцепления, т. e. локализацию генов в разных хромосомах. Это противоречие объясняется тем, что при составлении генетических карт суммируются расстояния между двумя наиболее близкими генами, что превышает экспериментально наблюдаемый процент кроссинговера.
Цитогенетическое картирование
Этот метод основан на использовании хромосомных перестроек. В случае гигантских политенных хромосом он позволяет прямо сопоставлять результаты генетического анализа расстояний между изучаемыми локусами и их взаимного расположения с данными о физических размерах определенных хромосомных областей. При облучении и действии других мутагенов в хромосомах часто наблюдаются выпадения (делеции) или вставки небольших фрагментов, сравнимых по величине с одним или несколькими локусами. Например, можно использовать гетерозиготы по хромосомам, одна из которых будет нести группу следующих друг за другом доминантных аллелей, тогда как гомологичная ей – группу рецессивных форм тех же генов. Если хромосома с доминантными генами будет последовательно терять отдельные локусы, то в гетерозиготе будут проявляться рецессивные признаки. Порядок проявления рецессивных признаков указывает на последовательность расположения генов.
При порядке генов AbC в случае делеции, захватывающей ген С, у мух с укороченной хромосомой, потерявшей фрагмент, равный гену С, в фенотипе проявятся аллели с, b и А.
В целом сравнение генетических (кроссинговерных) и цитологических карт показывает их соответствие: чем больший процент кроссинговера разделяет пару генов, тем больше и физическое расстояние между ними. Однако на несоответствие расстояний, определяемых указанными двумя методами, могут влиять два фактора. Во-первых, это области, в которых затруднен или отсутствует кроссинговер (например, в гетерохроматических районах); во-вторых, физическое расстояние будет больше, чем генетическое, если гены разделены зоной «молчащей» ДНК. Расчеты Бриджеса показали, что каждой единице перекреста на карте политенных хромосом слюнных желез D. melanogaster соответствует 4,2 мкм длины политенных хромосом. Эта длина как минимум равна двум-трем средним генам.
Особенности построения генетических карт у прокариот
Для построения генетических карт у прокариот используется явление конъюгации – переноса генетического материала из одной клетки в другую с помощью специальных кольцевых молекул ДНК (плазмид, в частности, с помощью F–плазмиды).
Вероятность переноса определенного гена в клетку–реципиент зависит от его удаления от F–плазмидной ДНК, а точнее, от точки О, в которой начинается репликация F–плазмидной ДНК. Чем больше время конъюгации, тем выше вероятность переноса данного гена. Это дает возможность составить генетическую карту бактерий в минутах конъюгации. Например, у кишечной палочки ген thr (оперон из трех генов, контролирующих биосинтез треонина) находится в нулевой точке (то есть непосредственно рядом с F–плазмидной ДНК), ген lac переносится через 8 мин, ген recE – через 30 мин, ген argR – через 70 мин и т.д.
Более подробно этот вопрос будет рассмотрен при изучении генетики прокариот.
Картирование хромосом человека
Картирование генов основано на составлении групп сцепления. Чем больше известных мутаций и чем меньше число хромосом, тем легче проводить картирование. В этом отношении человек (помимо того, что у него невозможен классический гибридологический анализ) как объект вдвойне неблагоприятен для картирования: известных генов у него сравнительно немного (по крайней мере, так было до конца 70-х годов), а гаплоидное число хромосом достаточно велико – 22 (не считая половых). Это означает, что вероятность того, что два вновь открытых гена окажутся сцепленными, равна 1/22. По этим причинам анализ родословных, который в какой-то мере заменяет гибридологический анализ, дает довольно ограниченную информацию о характере сцепления.
Более перспективными для картирования генов человека оказались методы генетики соматических клеток. Суть одного из них заключается в следующем. Методы клеточной инженерии позволяют объединять различные типы клеток. Слияние клеток, принадлежащих к разным биологическим видам, называется соматической гибридизацией. Сущность соматической гибридизации заключается в получении синтетических культур путем слияния протопластов различных видов организмов. Для слияния клеток используют различные физико-химические и биологические методы. После слияния протопластов образуются многоядерные гетерокариотические клетки. В дальнейшем при слиянии ядер образуются синкариотические клетки, содержащие в ядрах хромосомные наборы разных организмов. При делении таких клеток in vitro образуются гибридные клеточные культуры. В настоящее время получены и культивируются клеточные гибриды «человек × мышь», «человек × крыса» и многие другие.
В гибридных клетках, полученных из разных штаммов разных видов, один из родительских наборов хромосом, как правило, реплицируется быстрее другого. Поэтому последний постепенно теряет хромосомы. Эти процессы интенсивно протекают, например, в клеточных гибридах между мышью и человеком – видами, различающимися по многим биохимическим маркерам. Если при этом следить за каким-либо биохимическим маркером, например ферментом тимидинкиназой, и одновременно проводить цитогенетический контроль, идентифицируя хромосомы в клонах, образующихся после их частичной утраты, то, в конце концов, можно связать исчезновение хромосомы одновременно с биохимическим признаком. Это означает, что ген, кодирующий этот признак, локализован в данной хромосоме. Так, тимидинкиназный ген у человека находится в хромосоме 17.
Некоторая информация о локализации генов может быть получена при анализе числовых и структурных мутаций хромосом, по встречаемости в семьях хромосом с морфологическими вариациями и по учету наследственных признаков. Для этой же цели используют и частичные моносомии, возникающие в результате делеций. Однако в этих случаях необходимо иметь в виду, что иногда изучаемый ген остается в центрическом фрагменте, но его проявление может быть резко ослаблено в результате эффекта положения или каких-либо иных механизмов регуляции (изменение порядка репликации, отрыв промоторного участка и т. д.). В конце 60-х годов был разработан метод гибридизации in situ, в основе которого лежит специфичность комплементарных взаимодействий гена и его копии (мРНК, а также полученной с помощью обратной транскрипции комплементарной ДНК). Разрешающая способность этого метода гораздо выше на политенных хромосомах, чем на митотических хромосомах человека, однако он постоянно совершенствуется.
Генетические карты сцепления. Генетические карты сцепления определяют хромосомную принадлежность и взаимное расположение генетических маркеров относительно друг друга. Картирование в узком смысле -- определение положения гена или мутации в хромосоме. Позднее этот термин получил более широкое толкование. Он относится не только к гену, но к любому маркеру, под которым подразумевают ген, мутацию, участок ДНК с неопределенной функцией, точку расщепления ДНК рестриктазами. Таким образом, маркер -- это любой наследуемый признак, доступный идентификации тем или иным способом. Установление локализации какого-либо маркера позволяет использовать его для определения положения другого маркера.
На практике именно генетические карты сцепления и только они позволяют локализовать сложные генетические маркеры (например, ассоциированные с симптомами заболевания) на первых этапах исследования и дают возможность их дальнейшего изучения.
До начала 70-х годов XX в. построение генетических карт человека продвигалось очень медленными темпами. Первый ген человека (ген цветной слепоты) был картирован на Х-хромосоме в 1911 г., а первый аутосомный ген -- только в 1968 г. К 1973 г. на хромосомах человека было картировано 64 гена, а к 1994 г. -- 5000 структурных генов и свыше 60 000 маркерных ДНК-последовательностей. Столь стремительный прогресс в картировании генов человека связан с появлением новых технологий в цитогенети-ке, в клеточных культурах и особенно в молекулярной генетике.
Гибридизация соматических клеток. Одним из наиболее популярных методов отнесения генетического маркера (функционально активного гена) к конкретной группе сцепления является гибридизация (слияние друг с другом) соматических клеток разных биологических видов организмов, один из которых -- исследуемый. Гибридные клоны получают путем искусственного слияния клеток человека и различных грызунов: китайского хомячка, мыши, крысы. Культивирование таких соматических гибридов, как оказалось, сопровождается утратой хромосом человека. Потеря хромосом носит случайный характер, и образующиеся клоны клеток содержат оставшиеся хромосомы в разных сочетаниях. Так получают панели гибридных клеточных клонов, содержащих всего одну или несколько хромосом человека и полный набор хромосом другого вида. Обнаружение человеческих белков, специфических мРНК или последовательностей ДНК в таких клонах позволяет однозначно определить хромосомную принадлежность соответствующих генов.
Гибридизация in situ (в том же месте). Этот метод дает возможность локализовать определенные последовательности нуклеотидов на хромосомах. Они выступают в качестве зондов. Препараты фиксированных хромосом гибридизуют с исследуемыми последовательностями, меченными радиоактивной или флуоресцентной меткой. Меченые молекулы оказываются ассоциированными с участками хромосом, содержащими последовательности, комплементарные меченому зонду. Полученные гибриды анализируют с помощью микроскопа либо непосредственно, либо после радиоавтографии. Этот метод по частоте использования в последнее время прочно выходит на первое место. Наиболее популярной оказалась группа методов, получивших название флуоресцентной гибридизации in situ -- метод FISH (от англ. Fluorescence in situ hybridization ).
Полимеразная цепная реакция (ПЦР) позволила быстро и эффективно амплифицировать почти любой участок генома человека, а полученные продукты ПЦР использовать в качестве зондов для картирования соответствующих участков на хромосомах путем гибридизации in situ . В этом плане успешно разработана концепция сайтов, привязанных к последовательностям, --STS (от англ. Sequence-tagged sites). Все фрагменты ДНК, которые используются для построения генетических и физических карт, можно однозначно идентифицировать с помощью последовательности нуклеотидов длиной в 200 -- 500 н.п., которая является уникальной для данного фрагмента. Эти сайты амплифицируют с помощью ПЦР и применяют в качестве зондов. STS позволили создать основу для разработки единого языка, дающего возможность разным лабораториям описать свои клоны. Конечным результатом разработки концепции STS является создание исчерпывающей карты STS генома человека. Для получения маркеров в настоящее время часто применяют праймеры, соответствующие диспергированным повторяющимся последовательностям, среди которых первыми стали использовать А1u-последовательности, так как они характерны именно для генома человека. Поскольку в геноме человека больше 90 % умеренно повторяющихся последовательностей представлены семействами А1u и Крn I (последние повторяются реже и обладают характерной локализацией в хромосомах), они и используются для получения соответствующих зондов в ПЦР-реакции.
Физические карты низкого разрешения. Физические карты генома отражают реальное расстояние между маркерами, выражаемое в парах нуклеотидов. Физическую карту низкого разрешения часто называют хромосомной (цитогенетической) картой генома.
В начале 70-х годов XX в. появилась реальная возможность точной идентификации не только всех хромосом в кариотипе человека, но и их отдельных сегментов. Это связано с появлением мето да дифференциального окрашивания препаратов метафазных хромосом. Хромосомные препараты окрашивают некоторыми флуорохромами после соответствующей протеолитической обработки или нагревания. При этом на хромосомах выявляется характерная поперечная исчерченность -- так называемые диски (бэнды), расположение которых специфично для каждой хромосомы. Величина небольших дисков на прометафазных хромосомах соответствует примерно 1 млн н.п. на физических картах. Каждая хромосома после дифференциальной окраски может быть разделена на сегменты, нумерация которых начинается от центромерного района вверх (короткое плечо р) либо вниз (длинное плечо -- q) . Полосы в каждом сегменте также пронумерованы в аналогичном порядке. Запись положения гена на карте включает номер хромосомы, плечо, номер сегмента, бэнда и его субъединицы.
Запись 7 q21.1 означает, что ген локализован в субъединице 1-го бэнда 2-го сегмента длинного плеча хромосомы 7. Подобная запись удобна для цитогенетического картирования метода гибридизации in situ, позволяющего локализовать ген с точностью до одного бэнда и даже его субъединицы.
Хромосомные карты генома человека получают также локализацией генетических маркеров, чаще всего методом FISН: для метафазных хромосом разрешающая способность хромосомных карт находится в пределах 2 -- 5 млн н.п.; для интерфазных хромосом (генетический материал находится в менее компактной форме) -- приближается к 100 тыс. н.п. Для этого уровня картирования характерны карты кДНК (с. 358). Они отражают положение экспрес-сирующихся участков ДНК (экзонов) относительно известных ци-тогенетических маркеров (бэндов) на метафазных хромосомах. Поскольку такие карты дают представление о локализации транскрибирующихся участков генома, в том числе и генов с неизвестными функциями, они могут быть использованы для поиска новых генов. Этот подход полезен при поиске генов, повреждение которых вызывает заболевания человека, в том случае, если приблизительная локализация таких участков хромосом уже проведена на генетических картах сцепления (см. рис. 100).
Физические карты высокого разрешения. Для построения физических карт высокого разрешения экспериментально реализуется два альтернативных подхода: картирование сверху вниз и картирование снизу вверх (рис.В к геному) . Для картирования сверху вниз препарат ДНК индивидуальной хромосомы человека разрезают крупнощепящими рестриктазами (например, Not I) на длинные фрагменты, которые после разделения методом электрофореза в пульсирующем поле подвергаются дальнейшей обработке другими рестриктазами.
Методом электрофореза под действием однонаправленного постоянного поля в агарозном или полиакриламидном гелях удается разделить фрагменты ДНК размером не более 30 --50 тыс. н.п. Продвижение больших фрагментов ДНК в геле при пульсирующем изменении направления электрического поля происходит за счет конформационных изменений, обусловленных скручиванием и раскручиванием молекул ДНК в момент переключения направления поля. В этом случае удается разделить молекулы ДНК размером от 50 тыс. н.п. до 10 млн н.п.).
В результате получают макрорестрикционную карту. Метод электрофореза был с успехом использован для картирования малых геномов.
Для картирования генома человека снизу вверх на основе препарата суммарной ДНК генома или индивидуальной хромосомы получают серию случайных клонов протяженных последовательностей ДНК (10-- 1000 тыс. н.п.), часть из которых перекрывается друг с другом. В качестве вектора для клонирования в этом случае используют искусственные минихромосомы дрожжей (УАС). Последовательный набор клонов, содержащих частично перекрывающиеся и дополняющие друг друга фрагменты ДНК из определенного района генома, получил название скользящего зондирования, или «прогулки по хромосоме». Каждый раз отобранный фрагмент используется в качестве ДНК-зонда для последующего поиска. В результате получают набор клонированных фрагментов ДНК, полностью перекрывающих исследуемый участок генома, получивший название «контиг». Эта стратегия впервые была успешно применена для изучения 3-й хромосомы дрозофилы. С ее помощью редко удается пройти более 200 -- 300 тыс. н.п. в одном направлении из-за наличия в геноме повторяющихся и трудно клонируемых последовательностей ДНК. Для преодоления таких ограничений и ускорения процесса поиска генных последовательностей Ф. Коллинз, ныне президент Международного консорциума, предложил метод «прыжков» по хромосоме, позволяющий изолировать фрагменты ДНК, отстоящие в геноме друг от друга на сотни тысяч пар нуклеотидов (длина прыжка), не выделяя при этом все промежуточные последовательности ДНК.
Правильность полученных контигов подтверждают обычно гибридизацией in situ (FISH) с одновременной привязкой к определенным участкам исследуемых хромосом.
gene mapping, mapping - картирование генов.
Oпределение положения данного гена на какой-либо хромосоме относительно других генов; используют три основные группы методов К.г. - физическое (определение с помощью рестрикционных карт, электронной микроскопии и некоторых вариантов электрофореза межгенных расстояний - в нуклеотидах), генетическое (определение частот рекомбинаций между генами, в частности, в семейном анализе и др.) и цитогенетическое (гибридизации in situ <in situ hybridization >, получение монохромосомных клеточных гибридов <monochromosomal cell hybrid >, делеционный метод <deletion mapping > и др.); в генетике человека приняты 4 степени надежности локализации данного гена - подтвержденная (установлена в двух и более независимых лабораториях или на материале двух и более независимых тест-объектов), предварительная (1 лаборатория или 1 анализируемая семья), противоречивая (несовпадение данных разных исследователей), сомнительная (не уточненные окончательно данные одной лаборатории); в Приложении 5 приведена сводка (по состоянию на 1992-93) структурных генов, онкогенов и псевдогенов в геномах человека и - включая некоторые мутации - мыши.
(Источник: «Англо-русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд-во ВНИРО, 1995 г.)
- - составление схем, в к-рых гены расположены в линейном порядке с указанием относительных расстояний между ними. Методы Г. к.: комбинационные, рестрикционные, физические, транскрипционные, трансляционные...
Словарь микробиологии
- - изучение географического распределения аллелей генов с помощью картирования генетических данных по популяциям с последующим анализом геногеографических карт. Цель метода - изучение пространственных...
- - см. Географический метод...
Физическая Антропология. Иллюстрированный толковый словарь
- - genetic mapping - .Определение положения генов на генетической карте
, базирующееся на оценкe частоты рекомбинации между ними, а также маркерными генами... - - deletion mapping - .Mетод определения положения генных локусов в геноме с помощью небольших делеций
с известной локализацией... Молекулярная биология и генетика. Толковый словарь
- - denaturation mapping - .Метод идентификации частично денатурированных молекул ДНК по расположению вдоль нее легкоплавких АТ-богатых участков, которое отражает особенности первичной структуры...
Молекулярная биология и генетика. Толковый словарь
- - exclusive mapping - .Способ картирования генов на хромосоме
, состоящий в исключении участков, захватываемых делециями с известным положением... Молекулярная биология и генетика. Толковый словарь
- - backcross mapping - .Генетический метод картирования, основанный на получении бэккроссных гибридов родственных форм и анализе расщепления вариантов аллелей, полиморфных по длинам рестрикционных фрагментов <...
Молекулярная биология и генетика. Толковый словарь
- - crossing-over mapping - .Mетод определения межлокусных расстояний по частоте образования кроссоверных фенотипов, эти расстояния выражают в кроссоверных единицах; точность К.п.к. зависит от проявления интерференции <...
Молекулярная биология и генетика. Толковый словарь
- - positional cloning - .Начальный этап определения гена, мутация в котором обусловливает анализируемое НЗЧ...
Молекулярная биология и генетика. Толковый словарь
- - установление порядка расположения генов и относительного расстояния между ними в группе сцепления...
Большой медицинский словарь
- - теми, кто ввел это понятие, рассматривается как одна из геол. дисциплин, изучающая методы составления геол. карт и их практическое применение...
Геологическая энциклопедия
- - составление геоморфологической карты любого м-ба в поле в отличие от картографирования геоморфологического - составления обзорных сводных геоморфологических карт путем генерализации геоморфологических карт...
Геологическая энциклопедия
- - составление карт и картосхем биогеохим. районирования. Картирование биогеохимическое - необходимый метод изучения строения биосферы и районирования биогеохимического...
Экологический словарь
- - карт"...
Русский орфографический словарь
- - ...
Формы слова
"картирование генов" в книгах
1. Промоторы генов
Из книги Гены и развитие организма автора Нейфах Александр Александрович1. Промоторы генов В этом разделе мы кратко расскажем о том, какие нуклеотидные последовательности, прилегающие к генам, а иногда и внутри гена, ответственны за процесс транскрипции. У прокариот эти участки, с которыми связывается молекула РНК-полимеразы и откуда
Картирование генома человека
автора Глэд ДжонДупликация генов
Из книги Эволюция [Классические идеи в свете новых открытий] автора Марков Александр ВладимировичДупликация генов МНОГОФУНКЦИОНАЛЬНЫЕ ГЕНЫ - ОСНОВА ЭВОЛЮЦИОННЫХ НОВШЕСТВ.Мысль о том, что дупликация генов служит важнейшим источником эволюционных новшеств, была высказана еще в 1930-е годы выдающимся биологом Джоном Холдейном (Haldane, 1933). Сегодня в этом нет никаких
Картирование генома человека
Из книги Будущая эволюция человека. Евгеника XXI века автора Глэд ДжонКартирование генома человека Нам незачем богов напрасно беспокоить - Есть внутренности жертв, чтоб о войне гадать, Рабы, чтобы молчать, и камни, чтобы строить! Осип Мандельштам, «Природа - тот же Рим…» Генетика - молодая наука. Эволюция видов была по-настоящему открыта
Генов
Из книги Большая Советская Энциклопедия (ГЕ) автора БСЭКогнитивное картирование и операционное кодирование
Из книги Информационные войны [Основы военно-коммуникативных исследований] автора Почепцов Георгий ГеоргиевичКогнитивное картирование и операционное кодирование Теперь мы остановимся на когнитивном моделировании политики. Проблема когнитивного моделирования формулируется как поиск корреляции «между лингвистическими структурами текста и структурами представлений его
3 Интегральный интеллект: картирование паттернов человеческого улья
автора Хэмилтон Мэрилин3 Интегральный интеллект: картирование паттернов человеческого улья «Тот факт, что пчёлы сегодня располагают картой, необязательно значит, что у них она была до того, как эволюционировал их танец: карта могла прийти позднее в виде экономичного способа работы со сложной
Картирование основополагающих паттернов городской жизни
Из книги Интегральный город. Эволюционные интеллекты человеческого улья автора Хэмилтон МэрилинКартирование основополагающих паттернов городской жизни Для того чтобы понять природу городов как живых систем (и тем самым их фрактальную, голографическую и морфическую природу), мы должны рассмотреть четыре основополагающих карты городской жизни:? четырёхсекторную
Определение понятия «картирование идей»
автора Наст ДжеймиОпределение понятия «картирование идей» «Ментальная карта использует весь диапазон способностей коры головного мозга с применением слов, изображений, чисел, логики, ритма, цвета, а также пространственных отношений в единственной, уникально мощной технике. Благодаря
Препятствие 10: картирование идей в режиме реального времени
Из книги Эффект визуализации автора Наст ДжеймиПрепятствие 10: картирование идей в режиме реального времени Препятствие 10: «Я попытался картировать идеи во время трехчасового доклада. Я не мог определить, к чему ведет выступающий, и моя карта стала беспорядочной. Через 20 минут я сдался и снова принялся делать линейные
Глава 9 Командное картирование
Из книги Эффект визуализации автора Наст ДжеймиГлава 9 Командное картирование Одним из самых живых и нервных видов моей работы является инструктирование команд в процессе создания, совместного использования и объединения их идей по конкретному вопросу и определения приоритетности поставленных задач. Этот вид
Индивидуальное картирование
Из книги Эффект визуализации автора Наст ДжеймиИндивидуальное картирование Как только тема определена, каждый участник самостоятельно картирует свои идеи, где фиксирует все свои мысли на заданную тему. В случае проведения занятия по годовому стратегическому планированию я прошу участников фиксировать их мысли,
Картирование семинара в режиме реального времени – Лэндмарк форум
Из книги Эффект визуализации автора Наст ДжеймиКартирование семинара в режиме реального времени – Лэндмарк форум Чун Бу Лим занимал многие руководящие посты, а на данный момент является ведущим преподавателем Нджи Энн Политекник в Сингапуре. Он использует технику картирования идей с 2006 года и великолепно овладел
Глава 13 Заключительное задание: картирование идей в режиме реального времени
Из книги Эффект визуализации автора Наст ДжеймиГлава 13 Заключительное задание: картирование идей в режиме реального времени В этой главе мы рассмотрим: определение картирования идей в режиме реального времени; пять ключевых моментов для успешного картирования идей в режиме реального
Картирование коррупционных рисков и возможностей
Из книги автораКартирование коррупционных рисков и возможностей Коррупция проникает в таможенные органы множеством путей. Одним из крайних вариантов является политическая приватизация таможенной системы в целом, которая превращает таможню в инструмент политической элиты для